- EXCRÉTION (physiologie animale)
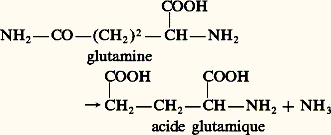
- EXCRÉTION (physiologie animale)L’excrétion représente un ensemble de processus de «triage», selon l’étymologie de ce vocable: moyens par lesquels des substances nocives ou inutiles sont éliminées hors de l’organisme, ou accumulées dans des tissus particuliers qui les mettent ainsi «hors circuit», autrement dit, en état d’exclusion. Quoique intuitive, cette notion de déchet est loin d’être simple: en effet, certaines substances, inutiles ou nocives pour la plupart des cellules animales, peuvent avoir, du fait de leur accumulation, une fonction physiologique, soit en jouant un rôle dans la coloration des animaux ou en les rendant impropres à la consommation, ce qui les protège des prédateurs éventuels (animaux vénéneux), soit en participant aux mécanismes de régulation fonctionnelle (cas du CO2). Les animaux tirent leur énergie de la dégradation de molécules organiques provenant de leurs aliments. Cette dégradation permet la formation de molécules vectrices d’énergie, tel l’ATP, et s’accompagne de la formation de composés résiduels. Les substances ne contenant que du carbone, de l’oxygène et de l’hydrogène (glucides et lipides) sont totalement dégradées en gaz carbonique (CO2) et en eau (H2O). Par contre, les substances qui contiennent en outre de l’azote (protéines et acides nucléiques) conduisent à un plus grand nombre de composés (urée, acide urique), variables selon les animaux. Selon la nature de ces substances, le problème de leur traitement par l’organisme peut se poser en termes différents. C’est pourquoi les fonctions d’excrétion peuvent être séparées en deux catégories bien distinctes: une fonction d’élimination des produits terminaux du métabolisme cellulaire et une fonction de régulation ou d’ajustement de la composition du milieu intérieur (eau, ions, diverses substances organiques dissoutes). Ce second volet ne sera pas envisagé ici, mais dans un autre contexte (cf. MILIEU INTÉRIEUR, OSMORÉGULATION, RESPIRATION).Compte tenu de la définition très large qui vient d’être donnée de l’excrétion, on peut attribuer à de nombreux organes une fonction excrétrice, qui est ou non leur principale fonction. Ainsi, chez l’Homme, en dehors des reins, qui éliminent de l’eau, des sels et diverses substances azotées, il convient de mentionner les poumons, qui rejettent du gaz carbonique et de l’eau, le foie, qui élabore la bile qu’il déverse ensuite dans le tube digestif (la bile contient de nombreuses substances organiques, comme des hormones inactivées ou divers composés toxiques d’origine exogène – médicaments, pesticides, etc.), ainsi que les glandes sudoripares, qui éliminent de l’eau, des sels et de l’urée. Notons que les divers organes ou glandes mentionnés possèdent d’autres fonctions: les poumons ont une fonction d’absorption, les glandes sudoripares une fonction thermorégulatrice, et le foie participe à de très nombreuses fonctions, métaboliques ou digestives. Pour ces raisons, seuls seront envisagés ici les organes dont la fonction excrétrice est la fonction principale.Après avoir envisagé la nature chimique des principaux déchets du catabolisme cellulaire, nous étudierons leur élimination. Celle-ci procède par deux mécanismes entièrement différents: soit leur accumulation dans certaines cellules spécialisées, soit leur rejet hors de l’organisme, grâce à des organes excréteurs (reins).Les produits terminaux du catabolisme azotéLes réactions du catabolisme cellulaire conduisent, d’une part, à la formation d’eau, de gaz carbonique et, d’autre part, à celle de divers composés azotés (fig. 1). Ceux-ci sont les seuls que l’on doive envisager dans le cadre de l’excrétion au sens strict. La dégradation des acides aminés et des bases pyrimidiques produit de l’ammoniac par des réactions de désamination (fig. 2). Ces réactions concernent environ 95 p. 100 de l’azote éliminé. Le reste, soit 5 p. 100, provient du catabolisme des bases puriques, qui conduit à l’acide urique. Ces réactions correspondent à un catabolisme primaire, qui ne permet pas à lui seul de rendre compte de la diversité des types d’excrétion azotée. Ainsi, l’Homme élimine environ 85 p. 100 de son azote en excès sous forme d’urée, les Oiseaux éliminent essentiellement de l’acide urique... ce qui n’est guère explicable par ce qui précède (qui correspondrait à 95 p. 100 d’ammoniac et 5 p. 100 d’acide urique). La grande diversité observée dans les formes d’excrétion azotée est due à l’existence de trois voies métaboliques supplémentaires (fig. 2):– la première correspond aux réactions de l’uricolyse , qui dégradent l’acide urique successivement en allantoïne, acide allantoïque, urée et, finalement, ammoniac;– la deuxième correspond à la formation d’urée , à partir de gaz carbonique et d’ammoniac (cycle de Krebs-Henselheit de l’uréogenèse), qui se produit principalement dans le foie chez les Vertébrés;– la troisième correspond aux réactions de néosynthèse purique , qui incorporent l’ammoniac dans des molécules d’hypoxanthine (précurseur des bases puriques). L’existence de ces trois voies, ou plutôt leur importance relative, permet de rendre compte de l’ensemble des cas observés, c’est-à-dire de la prédominance de tel ou tel composé azoté (ammoniac, urée ou acide urique).Suivant la nature du déchet azoté prédominant, on a défini un certain nombre de catégories d’animaux (cf. tableau). Ceux qui éliminent l’azote sous forme d’ammoniac sont dits ammoniotèles, ceux qui l’éliminent sous forme d’urée sont dits uréotèles, tandis que ceux qui excrètent de l’acide urique sont dits uricotèles. Il existe également des catégories intermédiaires, qui éliminent des quantités comparables de deux des trois composés précédents (exemple les uréo-uricotèles). Les Arachnides, enfin, se distinguent de l’ensemble en éliminant essentiellement de la guanine (guanotélisme). Il ne faudrait pas pour autant croire que les choses sont aussi simples. À côté des composés majeurs, il existe de nombreuses substances azotées dont la contribution reste modeste: acides aminés, créatine et créatinine, oxyde de triméthylamine... contenant 10 p. 100 ou plus de l’azote éliminé.Principales caractéristiques des divers déchets azotésLes divers déchets azotés présentent des propriétés très différentes, tant en ce qui concerne leur solubilité dans l’eau que leur toxicité. L’ammoniac est très soluble dans l’eau et est toxique, surtout chez les Vertébrés, où sa concentration ne doit pas excéder 1 mmole/litre. De fait, cette substance ne circule pas sous cette forme, mais sous une forme liée à l’acide glutamique (glutamine), et est libérée sous l’action de la glutaminase présente dans les organes excréteurs.
 L’urée est une substance neutre, très soluble et peu toxique, susceptible de s’accumuler dans le sang et les cellules à des concentrations variant de 10 à 300 mmoles/litre selon les espèces.L’acide urique et la guanine sont des composés très peu solubles dans l’eau (soit environ 0,17 mmole/litre pour l’acide urique à pH neutre), qui précipiteront très facilement sous forme de cristaux.En conséquence, on observe que l’ammoniac est le principal composé azoté excrété en milieu aquatique: cette molécule traverse facilement les membranes cellulaires et est très diffusible; elle est éliminée au niveau des téguments, des branchies, ainsi que des organes excréteurs sensu stricto . Ce mode d’excrétion azotée ne se retrouve pratiquement pas en milieu terrestre, sauf pour des animaux ayant un accès permanent à l’eau. En dehors de ces cas, les animaux terrestres éliminent soit de l’urée, soit de l’acide urique. L’urée, peu toxique, peut s’accumuler dans le sang et, par là même, se retrouver à de fortes concentrations dans l’urine ; elle nécessite donc beaucoup moins d’eau pour son élimination qu’une quantité égale d’ammoniac. L’acide urique, quant à lui, est susceptible d’être éliminé sous forme pâteuse, presque sans eau: on le trouve chez les animaux pour lesquels l’économie de l’eau est un problème majeur (Insectes, Reptiles), ainsi que chez les Oiseaux, où ce mode d’excrétion permet de réduire les quantités d’eau emmagasinée en vue de l’élimination des déchets azotés, donc le poids. Par ailleurs, l’acide urique et la guanine, de par leur très faible solubilité, se prêtent aisément à un stockage intracellulaire sous forme cristalline (par exemple, dans le corps gras des Insectes).La relation étroite entre le milieu où vivent les animaux et la nature des composés azotés excrétés est particulièrement bien illustrée avec le cas des Amphibiens: le têtard, aquatique, est ammoniotélique, tandis que la grenouille, terrestre, est uréotélique. Le mode de vie peut également moduler l’excrétion: ainsi, tandis que la plupart des Oiseaux terrestres éliminent plus de 90 p. 100 de l’azote sous forme d’acide urique, les canards, qui mènent une vie semi-aquatique, éliminent 30 p. 100 de l’azote sous forme d’ammoniac (sels d’ammonium). De même, les Reptiles peuvent être strictement uricotéliques (lézards), uréo-uricotéliques (tortues), ou ammonio-uricotéliques (crocodiles). Le mode d’excrétion d’un même individu n’est pas nécessairement fixe: lorsque les tortues sont soumises à des conditions de sécheresse, elles deviennent totalement uricotéliques. Les variations peuvent être saisonnières: ainsi, le Gastéropode Pila globosa est ammoniotélique pendant sa période de vie active (aquatique), et devient uricotélique pendant sa période d’hibernation en milieu terrestre (ce qui lui permet d’accumuler les déchets azotés sous une forme cristallisée – osmotiquement inactive).Ces variations adaptatives ont toutefois des limites et, au cours de l’évolution des divers groupes animaux, certaines orientations ont été fixées de manière irréversible. Ainsi, les Mammifères marins, qui sont issus de formes terrestres, sont uréotèles et ne sont pas devenus ammoniotèles. De même, certains Gastéropodes d’eau douce (limnées) sont uricotèles, comme les espèces dont ils dérivent.Il faut enfin noter que d’autres pressions de sélection ont conduit à l’existence d’animaux marins uréotèles: les Sélaciens (requins, raies). Ces animaux sont issus de formes d’eau douce ammoniotèles. L’urée qu’ils forment (ainsi que l’oxyde de triméthylamine – essentiellement d’origine alimentaire) s’accumule dans le sang et les tissus, et permet à ces animaux de résoudre les problèmes d’osmorégulation de manière originale.L’excrétion par accumulation dans l’organismeLes déchets du métabolisme ne sont pas toujours éliminés hors de l’organisme. Certains d’entre eux, et plus particulièrement les moins hydrosolubles, peuvent être stockés de façon temporaire ou permanente dans des cellules ou des organes particuliers, ceci ne résultant pas nécessairement d’une incapacité à éliminer les substances en question.L’excrétion par accumulation est en fait un processus fréquemment rencontré chez les animaux. Les auteurs du siècle dernier ont à ce sujet effectué une recherche systématique, qui consistait souvent à injecter à des animaux de l’encre de Chine (fine suspension de granules de mélanine). En suivant le devenir de ces particules, ils constatèrent qu’elles s’accumulaient dans des cellules particulières, mobiles ou fixes, isolées ou groupées en tissus. Cette accumulation pouvait être permanente, les cellules ainsi chargées en pigments n’évoluant plus, ou transitoire, les cellules se désagrégeant par migration hors de l’animal, ou transférant leur contenu dans des sites d’accumulation permanente.Dans la pratique, de tels systèmes ont été particulièrement étudiés chez les Invertébrés, parmi lesquels nous retiendrons quelques exemples. Chez les Annélides terrestres, notamment le lombric, la paroi des vaisseaux sanguins et du tube digestif est bordée par des chloragocytes (fig. 3), qui se chargent de déchets formant des inclusions nombreuses dans les cellules (concréments). Ces cellules représentent un site d’accumulation renouvelable, les déchets étant finalement stockés dans l’épiderme et les cellules sous-jacentes. Les chloragocytes sont particulièrement développés chez les sangsues, où ils forment le tissu botryoïdal, riche en pigments biliaires provenant de la dégradation de l’hémoglobine ingérée par ces animaux hématophages (l’épiderme contient également de grandes quantités de ces pigments).Chez les Insectes, le rôle de «rein d’accumulation» est tenu par les cellules épidermiques – les belles couleurs des ailes des papillons sont ainsi à mettre en relation (en partie du moins) avec un rôle de rein que jouent les ailes au cours de la vie nymphale – et par le corps gras, équivalent du foie des Vertébrés, qui accumule quant à lui des quantités notables d’acide urique: chez certaines blattes, l’acide urique ainsi stocké peut représenter jusqu’à 10 p. 100 du poids total du corps!Chez les Vertébrés, on connaît des exemples similaires. La chair des saumons est colorée en rose par des pigments d’origine alimentaire, et l’usage des caroténoïdes alimentaires permet aussi bien d’obtenir des poulets à pattes jaunes (nourris au maïs), des canaris au plumage resplendissant, que de donner une apparence bronzée à la peau humaine (faux bronzage, car ne faisant pas intervenir les mêmes pigments). Ces derniers exemples sont tout à fait comparables à ce que l’on a vu chez les Invertébrés, et soulignent le rôle de la peau et de ses productions (plumes, écailles, etc.). Le foie des Vertébrés possède une fonction similaire. Cet organe joue un rôle de premier plan dans les mécanismes de détoxification. Il est normalement chargé de dégrader les substances indésirables (endogènes ou exogènes) et de les transformer en des formes éliminables par les reins, ou de les éliminer directement dans la bile. La capacité de détoxification du foie a toutefois des limites, et le foie est alors susceptible d’accumuler certaines substances: médicaments, pesticides... sont des exemples classiques de composés s’accumulant dans le foie des animaux.L’accumulation des déchets peut prendre une forme très particulière chez certains Tuniciers (ascidies appartenant à la famille des Molgulidés). Ces animaux possèdent un rein qui est constitué par une vésicule close qui accumule des concrétions riches en acide urique (toutefois, la majorité de l’azote est éliminée sous forme d’ammoniac). Dans ce cas, le stockage est extracellulaire, et peut être rapproché d’un stockage de déchets se produisant dans la cuticule des Insectes.L’excrétion par accumulation ne représente toutefois pas l’essentiel de l’élimination, la plus importante étant l’élimination hors de l’animal.L’excrétion par rejet hors de l’organismeLes animaux unicellulaires représentent un premier niveau d’organisation, selon lequel l’excrétion procède en général à travers la membrane cellulaire. Certains d’entre eux possèdent toutefois des structures spécialisées, dont la fonction principale est sans doute osmorégulatrice. C’est le cas des Protozoaires ciliés (paramécies), qui possèdent des vacuoles pulsatiles éliminant l’excès d’eau [cf. PROTISTES].Les Métazoaires les plus primitifs (Porifères - épongesa - et Cœlentérés), ainsi que quelques groupes plus évolués (Échinodermes, Urocordés, qui disposent de reins d’accumulation), sont dépourvus d’organes excréteurs spécialisés; ils rejettent donc les déchets à travers leur tégument.Les autres animaux possèdent des organes excréteurs plus ou moins complexes, grâce auxquels ils rejettent les déchets à l’extérieur du corps. Ces organes sont constitués par des unités appelées néphridies chez les Invertébrés et néphrons chez les Vertébrés. Ces unités peuvent rester isolées, ou se regrouper en structures plus complexes qui portent alors le nom de reins . Les aspects que peuvent prendre ces organes sont très variables, mais ils présentent cependant une grande unité en ce qui concerne les mécanismes cellulaires de leur fonctionnement.Principes fonctionnels générauxOn retrouve de manière constante trois processus de base (fig. 4) dans le fonctionnement des organes excréteurs, qui sont la filtration , qui permet la formation d’une urine primaire, suivie de processus d’élaboration d’une urine définitive par le jeu de réabsorptions et de sécrétions .La filtration peut faire intervenir trois modalités différentes. Dans le premier cas, il existe une pression de filtration liée à l’existence d’une pression relativement importante dans le système circulatoire, qui tend à faire sortir les liquides circulants à l’extérieur des vaisseaux. Cela est en particulier réalisé au niveau des glomérules des néphrons de Vertébrés, ainsi qu’au niveau de divers vaisseaux sanguins ou des parties contractiles du système circulatoire chez divers Invertébrés (Mollusques, Crustacés, etc.). Les parois des capillaires (Vertébrés) ou de l’appareil circulatoire sont alors doublées par des cellules particulières, les podocytes, et ce système réalise une ultrafiltration du sang ou de l’hémolymphe. Cela signifie que l’urine primitive ainsi formée a une composition très proche de celle des fluides circulants (seules les grosses molécules comme les protéines sont arrêtées par ce filtre). Chez les Invertébrés où le cœlome est développé (ainsi que chez les embryons de Vertébrés), l’ultrafiltration alimente le liquide cœlomique, qui contient également des cellules circulantes (cœlomocytes). L’élimination de l’urine s’effectue alors par une métanéphridie ouverte.Un deuxième mode de filtration est rencontré dans le cas des protonéphridies. Ces structures tubulaires plus ou moins ramifiées possèdent à une extrémité des cellules particulières, appelées cirtocytes ou solénocytes. Les cirtocytes possèdent une flamme vibratile formée par une touffe de cils agglutinés; les solénocytes possèdent un seul flagelle. Les battements des cils ou du flagelle, mettant en mouvement le fluide intratubulaire, créent une dépression dans la partie terminale (fig. 5), ce qui entraîne une ultrafiltration des liquides interstitiels (Acœlomates) ou du liquide cœlomique (Cœlomates). Le résultat final se rapproche de celui du premier cas, mais appliqué à des animaux dépourvus d’appareil circulatoire élaboré.Un troisième mécanisme se rencontre dans le cas des néphrons aglomérulés (Téléostéens marins: l’hippocampe par exemple) et dans le cas des tubes de Malpighi des Insectes. Ces organes ont des origines embryologiques différentes, mais fonctionnent selon le même principe. Il n’existe pas d’appareil filtrant, et le mécanisme primaire est un transport actif d’ions (ions bivalents chez les Poissons, potassium chez les Insectes), qui entraîne un flux d’eau passif. La composition de l’urine primaire est alors très différente de celle du plasma ou de l’hémolymphe (elle est ainsi très enrichie en potassium chez les Insectes).Dans l’ensemble des cas considérés, l’urine éliminée est très différente de l’urine primaire. Cette modification est liée à des processus de sécrétion (NH3, par exemple) et de réabsorption tubulaires ou se produisant dans des parties extra-rénales: vessie des Amphibiens, cloaque des Oiseaux, rectum des Insectes...Protonéphridies et métanéphridiesLes protonéphridies apparaissent chez les Platheminthes (planaires). Elles dérivent dans ce cas de l’ectoderme, mais cela n’est pas une règle générale. Elles sont constituées par un ensemble de cellules qui s’ouvrent sur un canal excréteur. Cette structure peut être unique, paire ou multiple, à disposition métamérique (Annélides). Les protonéphridies, comme de nombreux organes excréteurs, ont un double rôle: élimination des déchets du métabolisme et régulation osmotique-ionique. La vitesse de filtration (de formation de l’urine primaire) est directement liée à la fréquence des battements de la flamme ou des flagelles.Les métanéphridies sont d’origine mésodermique. Elles sont ouvertes à leurs deux extrémités. L’ouverture interne, dans le cœlome (néphridiostome, ou pavillon cilié), réalise une filtration et non une ultrafiltration (celle-ci se produit entre le système circulatoire et le cœlome). Ce système est anatomiquement très comparable aux néphrons ouverts des embryons des Vertébrés, qui possèdent des glomérules intracœlomiques. La structure des métanéphridies est en fait complexe, et le tubule comporte plusieurs régions spécialisées dans la sécrétion ou la réabsorption (fig. 6). Ces échanges se produisent avec des vaisseaux sanguins qui longent le tubule.Chez les Annélides, les métanéphridies sont nombreuses: une paire par métamère. Chez les Mollusques, au contraire, le cœlome est réduit au cœlome péricardique et périgonadique, et l’on trouve soit une paire de métanéphridies ouvrant dans le péricarde, soit même une seule métanéphridie, comme dans le cas des Gastéropodes pulmonés où elle porte le nom d’«organe de Bojanus». La structure en est complexe (fig. 7). L’urine primaire se forme par ultrafiltration au niveau de la paroi cardiaque et de la partie proximale de la néphridie, richement vascularisée, qui porte le nom de «rein». L’épithélium de ce rein est très plissé, et sécrète en outre des grains de concrétions riches en urates et en diverses purines. L’urine finale est hypo-osmotique, en raison d’une importante réabsorption de sels dans la partie distale de la néphridie (uretère). On voit là de grandes analogies avec les Vertébrés, analogies qui sont soulignées par l’emploi des mêmes termes (rein, uretère). Chez les Mollusques céphalopodes (marins), l’épithélium néphridien sécrète des ions ammonium et non des urates, mais, malgré une anatomie quelque peu différente, le fonctionnement est très comparable.Chez les Crustacés (écrevisse), on trouve une paire de reins céphaliques. Il s’agit, là encore, de métanéphridies où la réduction du cœlome rapproche considérablement le vaisseau sanguin afférent (artère antennaire) de la partie proximale de la néphridie, ce qui conduit à une structure analogue à celle d’un néphron glomérulé.Les tubes de Malpighi des InsectesL’appareil excréteur des Insectes comporte un nombre variable de tubes excréteurs, ou tubes de Malpighi, qui sont aveugles à leur extrémité distale et qui débouchent à la limite entre l’intestin moyen et l’intestin postérieur. Les tubes de Malpighi baignent dans l’hémolymphe. La formation de l’urine est liée à un transport actif d’ions potassium par les cellules de la partie distale: il n’y a ici ni filtration, ni ultrafiltration. Le flux d’ions potassium entraîne un flux d’eau passif. Diverses substances, présentes en solution dans l’hémolymphe, diffusent ensuite passivement vers la lumière du tubule, selon le gradient de concentration. Certaines d’entre elles, en particulier l’acide urique, pourraient faire l’objet d’un transport actif. L’urine primitive ainsi formée est sensiblement iso-osmotique à l’hémolymphe, bien que sa composition soit notablement différente, car elle est très enrichie en ions potassium. Elle est neutre ou légèrement alcaline, l’acide urique y est présent sous forme d’urate de potassium, relativement soluble.L’appareil excréteur est en fait constitué par l’ensemble tubes de Malpighi + intestin postérieur (iléon + rectum). Au niveau de l’intestin, l’urine est acidifiée, ce qui précipite les urates sous la forme d’acide urique; les ions et l’eau sont en grande partie réabsorbés, de même que de petites molécules organiques solubles. En fait, chez de nombreuses espèces, la réabsorption commence dans la partie proximale des tubes de Malpighi. Le système excréteur peut présenter diverses adaptations, permettant en particulier une plus grande efficacité dans la réabsorption de l’eau (système cryptonéphridien des chenilles de Lépidoptères, par exemple).Les néphrons des VertébrésLes reins des Vertébrés sont constitués de néphrons (fig. 4 a), qui comportent en général deux parties: le corpuscule de Malpighi (ensemble glomérule + capsule de Bowman) et le tubule, où s’effectuent réabsorptions et sécrétions. Ce schéma correspond à la majorité des Vertébrés adultes, mais on connaît deux variantes, la première correspondant aux néphrons ouverts , analogues à des métanéphridies, que l’on rencontre au cours du développement embryonnaire des Vertébrés inférieurs, la seconde aux néphrons aglomérulés de certaines espèces de Poissons téléostéens marins, qui fonctionnent par sécrétion (transport actif d’ions bivalents Mg++, Ca++, S4—, HP4—), à la manière des tubes de Malpighi des Insectes (fig. 4 c). Le tubule comprend une partie proximale (réabsorption obligatoire, sécrétion) et une partie distale (réabsorption facultative modulable), qui sont séparées par un segment intermédiaire, essentiellement développé chez les Oiseaux et surtout les Mammifères, où il est recourbé en U et forme l’anse de Henlé , à laquelle est liée l’aptitude à produire une urine hyperosmotique. Chez les autres Vertébrés, l’urine est soit iso-osmotique, soit hypo-osmotique.Les reins sont formés d’un nombre très variable de néphrons selon les espèces: de quelques dizaines à quelques millions – deux millions chez l’Homme. Leur mise en place s’effectue au cours du développement embryonnaire, en concernant des métamères de plus en plus distaux. On note successivement la formation d’un pronéphros , en position antérieure, qui n’est fonctionnel que chez les embryons des Vertébrés inférieurs, puis d’un mésonéphros , qui forme le rein des Poissons et des Amphibiens, enfin d’un métanéphros , qui constitue le rein des Vertébrés amniotes (Reptiles, Oiseaux et Mammifères).L’urine émise par les reins transite par les uretères et est souvent stockée transitoirement dans une vessie. Chez les Poissons téléostéens, les Amphibiens et les Reptiles chéloniens, cet organe peut jouer un rôle important, en réabsorbant de l’eau et diverses substances dissoutes. Chez les autres Reptiles et chez les Oiseaux, les voies urinaires rejoignent les voies digestives au niveau du cloaque, et l’urine remonte dans le colon. Dans ce cas, la muqueuse intestinale peut réabsorber l’eau et les sels, modifier le pH et accroître la précipitation de l’acide urique à la périphérie des fèces, ce qui leur donne leur couleur blanchâtre. Ce dernier cas est fonctionnellement très proche de l’ensemble tubes de Malpighi + rectum des Insectes.Les Mammifères, grâce au développement de l’anse de Henlé qui permet la formation d’une urine hyper-osmotique, possèdent la faculté de concentrer l’urine plus ou moins fortement selon les espèces, cela étant en relation avec le mode de vie: les espèces marines (baleines, phoques), ainsi que celles vivant en milieu désertique (rat-kangourou, dromadaire) présentent une aptitude maximale à cette fonction, liée à la très grande longueur des anses de Henlé (cf. vie dans les DÉSERTS).Le problème de l’élimination des sels en excès se pose également chez d’autres Vertébrés vivant en milieu marin ou désertique (Oiseaux, comme les goélands, Reptiles: iguanes, tortues, etc.), qui ne possèdent pas cette aptitude à concentrer l’urine (absence ou faible développement de l’anse de Henlé). Dans ce cas, les sels en excès sont éliminés par des organes spécialisés, les glandes à sels, capables de sécréter des solutions concentrées de chlorure de sodium ou de potassium.Les activités rénales sont contrôlées à divers niveaux, celui de la filtration glomérulaire, celui des transferts tubulaires. Il faut toutefois noter que ces contrôles sont plutôt à mettre en relation avec les problèmes d’osmorégulation que ceux d’excrétion (cf. OSMORÉGULATION, métabolisme HYDROMINÉRAL).Quelques aspects évolutifs de la fonction excrétriceAu cours de l’évolution animale, on rencontre successivement des protonéphridies, puis des métanéphridies et, enfin, des néphrons glomérulés, dont nous avons vu les similitudes de fonctionnement.Cette évolution se retrouve au cours du développement embryonnaire ou larvaire: ainsi, les larves trocophores des Mollusques possèdent des protonéphridies à solénocytes ou à cirtocytes, les adultes des métanéphridies; les embryons de Poissons possèdent des néphrons ouverts (équivalant à des métanéphridies), les adultes des néphrons fermés classiques.Des systèmes homologues peuvent présenter des adaptations convergentes: les larves de certains Mollusques marins possèdent des protonéphridies dépourvues de solénocytes ou cirtocytes, ce qui peut être rapproché du cas des néphrons aglomérulés de certains téléostéens marins.Les similitudes physiologiques s’accompagnent de similitudes biochimiques concernant la nature des principaux déchets azotés, qui ne dépend pas de la position systématique des animaux, mais du milieu dans lequel ils vivent. Les produits terminaux du métabolisme azoté ne sont pas nécessairement uniquement des déchets. Outre un rôle dans la pigmentation, signalé plus haut, ils peuvent avoir un rôle dans l’osmorégulation. C’est ainsi que l’urée, en s’accumulant dans le sang (et les cellules) sert à rendre les Sélaciens iso-osmotiques à l’eau de mer, ou à empêcher la dessication des Dipneustes et des Amphibiens lors de périodes d’estivation pendant lesquelles ils sont enfouis dans de la boue (dont la pression osmotique s’accroît au cours de l’assèchement). Ce même composé contribue également à l’économie de l’eau chez les Mammifères, en participant aux mécanismes de concentration urinaire.
L’urée est une substance neutre, très soluble et peu toxique, susceptible de s’accumuler dans le sang et les cellules à des concentrations variant de 10 à 300 mmoles/litre selon les espèces.L’acide urique et la guanine sont des composés très peu solubles dans l’eau (soit environ 0,17 mmole/litre pour l’acide urique à pH neutre), qui précipiteront très facilement sous forme de cristaux.En conséquence, on observe que l’ammoniac est le principal composé azoté excrété en milieu aquatique: cette molécule traverse facilement les membranes cellulaires et est très diffusible; elle est éliminée au niveau des téguments, des branchies, ainsi que des organes excréteurs sensu stricto . Ce mode d’excrétion azotée ne se retrouve pratiquement pas en milieu terrestre, sauf pour des animaux ayant un accès permanent à l’eau. En dehors de ces cas, les animaux terrestres éliminent soit de l’urée, soit de l’acide urique. L’urée, peu toxique, peut s’accumuler dans le sang et, par là même, se retrouver à de fortes concentrations dans l’urine ; elle nécessite donc beaucoup moins d’eau pour son élimination qu’une quantité égale d’ammoniac. L’acide urique, quant à lui, est susceptible d’être éliminé sous forme pâteuse, presque sans eau: on le trouve chez les animaux pour lesquels l’économie de l’eau est un problème majeur (Insectes, Reptiles), ainsi que chez les Oiseaux, où ce mode d’excrétion permet de réduire les quantités d’eau emmagasinée en vue de l’élimination des déchets azotés, donc le poids. Par ailleurs, l’acide urique et la guanine, de par leur très faible solubilité, se prêtent aisément à un stockage intracellulaire sous forme cristalline (par exemple, dans le corps gras des Insectes).La relation étroite entre le milieu où vivent les animaux et la nature des composés azotés excrétés est particulièrement bien illustrée avec le cas des Amphibiens: le têtard, aquatique, est ammoniotélique, tandis que la grenouille, terrestre, est uréotélique. Le mode de vie peut également moduler l’excrétion: ainsi, tandis que la plupart des Oiseaux terrestres éliminent plus de 90 p. 100 de l’azote sous forme d’acide urique, les canards, qui mènent une vie semi-aquatique, éliminent 30 p. 100 de l’azote sous forme d’ammoniac (sels d’ammonium). De même, les Reptiles peuvent être strictement uricotéliques (lézards), uréo-uricotéliques (tortues), ou ammonio-uricotéliques (crocodiles). Le mode d’excrétion d’un même individu n’est pas nécessairement fixe: lorsque les tortues sont soumises à des conditions de sécheresse, elles deviennent totalement uricotéliques. Les variations peuvent être saisonnières: ainsi, le Gastéropode Pila globosa est ammoniotélique pendant sa période de vie active (aquatique), et devient uricotélique pendant sa période d’hibernation en milieu terrestre (ce qui lui permet d’accumuler les déchets azotés sous une forme cristallisée – osmotiquement inactive).Ces variations adaptatives ont toutefois des limites et, au cours de l’évolution des divers groupes animaux, certaines orientations ont été fixées de manière irréversible. Ainsi, les Mammifères marins, qui sont issus de formes terrestres, sont uréotèles et ne sont pas devenus ammoniotèles. De même, certains Gastéropodes d’eau douce (limnées) sont uricotèles, comme les espèces dont ils dérivent.Il faut enfin noter que d’autres pressions de sélection ont conduit à l’existence d’animaux marins uréotèles: les Sélaciens (requins, raies). Ces animaux sont issus de formes d’eau douce ammoniotèles. L’urée qu’ils forment (ainsi que l’oxyde de triméthylamine – essentiellement d’origine alimentaire) s’accumule dans le sang et les tissus, et permet à ces animaux de résoudre les problèmes d’osmorégulation de manière originale.L’excrétion par accumulation dans l’organismeLes déchets du métabolisme ne sont pas toujours éliminés hors de l’organisme. Certains d’entre eux, et plus particulièrement les moins hydrosolubles, peuvent être stockés de façon temporaire ou permanente dans des cellules ou des organes particuliers, ceci ne résultant pas nécessairement d’une incapacité à éliminer les substances en question.L’excrétion par accumulation est en fait un processus fréquemment rencontré chez les animaux. Les auteurs du siècle dernier ont à ce sujet effectué une recherche systématique, qui consistait souvent à injecter à des animaux de l’encre de Chine (fine suspension de granules de mélanine). En suivant le devenir de ces particules, ils constatèrent qu’elles s’accumulaient dans des cellules particulières, mobiles ou fixes, isolées ou groupées en tissus. Cette accumulation pouvait être permanente, les cellules ainsi chargées en pigments n’évoluant plus, ou transitoire, les cellules se désagrégeant par migration hors de l’animal, ou transférant leur contenu dans des sites d’accumulation permanente.Dans la pratique, de tels systèmes ont été particulièrement étudiés chez les Invertébrés, parmi lesquels nous retiendrons quelques exemples. Chez les Annélides terrestres, notamment le lombric, la paroi des vaisseaux sanguins et du tube digestif est bordée par des chloragocytes (fig. 3), qui se chargent de déchets formant des inclusions nombreuses dans les cellules (concréments). Ces cellules représentent un site d’accumulation renouvelable, les déchets étant finalement stockés dans l’épiderme et les cellules sous-jacentes. Les chloragocytes sont particulièrement développés chez les sangsues, où ils forment le tissu botryoïdal, riche en pigments biliaires provenant de la dégradation de l’hémoglobine ingérée par ces animaux hématophages (l’épiderme contient également de grandes quantités de ces pigments).Chez les Insectes, le rôle de «rein d’accumulation» est tenu par les cellules épidermiques – les belles couleurs des ailes des papillons sont ainsi à mettre en relation (en partie du moins) avec un rôle de rein que jouent les ailes au cours de la vie nymphale – et par le corps gras, équivalent du foie des Vertébrés, qui accumule quant à lui des quantités notables d’acide urique: chez certaines blattes, l’acide urique ainsi stocké peut représenter jusqu’à 10 p. 100 du poids total du corps!Chez les Vertébrés, on connaît des exemples similaires. La chair des saumons est colorée en rose par des pigments d’origine alimentaire, et l’usage des caroténoïdes alimentaires permet aussi bien d’obtenir des poulets à pattes jaunes (nourris au maïs), des canaris au plumage resplendissant, que de donner une apparence bronzée à la peau humaine (faux bronzage, car ne faisant pas intervenir les mêmes pigments). Ces derniers exemples sont tout à fait comparables à ce que l’on a vu chez les Invertébrés, et soulignent le rôle de la peau et de ses productions (plumes, écailles, etc.). Le foie des Vertébrés possède une fonction similaire. Cet organe joue un rôle de premier plan dans les mécanismes de détoxification. Il est normalement chargé de dégrader les substances indésirables (endogènes ou exogènes) et de les transformer en des formes éliminables par les reins, ou de les éliminer directement dans la bile. La capacité de détoxification du foie a toutefois des limites, et le foie est alors susceptible d’accumuler certaines substances: médicaments, pesticides... sont des exemples classiques de composés s’accumulant dans le foie des animaux.L’accumulation des déchets peut prendre une forme très particulière chez certains Tuniciers (ascidies appartenant à la famille des Molgulidés). Ces animaux possèdent un rein qui est constitué par une vésicule close qui accumule des concrétions riches en acide urique (toutefois, la majorité de l’azote est éliminée sous forme d’ammoniac). Dans ce cas, le stockage est extracellulaire, et peut être rapproché d’un stockage de déchets se produisant dans la cuticule des Insectes.L’excrétion par accumulation ne représente toutefois pas l’essentiel de l’élimination, la plus importante étant l’élimination hors de l’animal.L’excrétion par rejet hors de l’organismeLes animaux unicellulaires représentent un premier niveau d’organisation, selon lequel l’excrétion procède en général à travers la membrane cellulaire. Certains d’entre eux possèdent toutefois des structures spécialisées, dont la fonction principale est sans doute osmorégulatrice. C’est le cas des Protozoaires ciliés (paramécies), qui possèdent des vacuoles pulsatiles éliminant l’excès d’eau [cf. PROTISTES].Les Métazoaires les plus primitifs (Porifères - épongesa - et Cœlentérés), ainsi que quelques groupes plus évolués (Échinodermes, Urocordés, qui disposent de reins d’accumulation), sont dépourvus d’organes excréteurs spécialisés; ils rejettent donc les déchets à travers leur tégument.Les autres animaux possèdent des organes excréteurs plus ou moins complexes, grâce auxquels ils rejettent les déchets à l’extérieur du corps. Ces organes sont constitués par des unités appelées néphridies chez les Invertébrés et néphrons chez les Vertébrés. Ces unités peuvent rester isolées, ou se regrouper en structures plus complexes qui portent alors le nom de reins . Les aspects que peuvent prendre ces organes sont très variables, mais ils présentent cependant une grande unité en ce qui concerne les mécanismes cellulaires de leur fonctionnement.Principes fonctionnels générauxOn retrouve de manière constante trois processus de base (fig. 4) dans le fonctionnement des organes excréteurs, qui sont la filtration , qui permet la formation d’une urine primaire, suivie de processus d’élaboration d’une urine définitive par le jeu de réabsorptions et de sécrétions .La filtration peut faire intervenir trois modalités différentes. Dans le premier cas, il existe une pression de filtration liée à l’existence d’une pression relativement importante dans le système circulatoire, qui tend à faire sortir les liquides circulants à l’extérieur des vaisseaux. Cela est en particulier réalisé au niveau des glomérules des néphrons de Vertébrés, ainsi qu’au niveau de divers vaisseaux sanguins ou des parties contractiles du système circulatoire chez divers Invertébrés (Mollusques, Crustacés, etc.). Les parois des capillaires (Vertébrés) ou de l’appareil circulatoire sont alors doublées par des cellules particulières, les podocytes, et ce système réalise une ultrafiltration du sang ou de l’hémolymphe. Cela signifie que l’urine primitive ainsi formée a une composition très proche de celle des fluides circulants (seules les grosses molécules comme les protéines sont arrêtées par ce filtre). Chez les Invertébrés où le cœlome est développé (ainsi que chez les embryons de Vertébrés), l’ultrafiltration alimente le liquide cœlomique, qui contient également des cellules circulantes (cœlomocytes). L’élimination de l’urine s’effectue alors par une métanéphridie ouverte.Un deuxième mode de filtration est rencontré dans le cas des protonéphridies. Ces structures tubulaires plus ou moins ramifiées possèdent à une extrémité des cellules particulières, appelées cirtocytes ou solénocytes. Les cirtocytes possèdent une flamme vibratile formée par une touffe de cils agglutinés; les solénocytes possèdent un seul flagelle. Les battements des cils ou du flagelle, mettant en mouvement le fluide intratubulaire, créent une dépression dans la partie terminale (fig. 5), ce qui entraîne une ultrafiltration des liquides interstitiels (Acœlomates) ou du liquide cœlomique (Cœlomates). Le résultat final se rapproche de celui du premier cas, mais appliqué à des animaux dépourvus d’appareil circulatoire élaboré.Un troisième mécanisme se rencontre dans le cas des néphrons aglomérulés (Téléostéens marins: l’hippocampe par exemple) et dans le cas des tubes de Malpighi des Insectes. Ces organes ont des origines embryologiques différentes, mais fonctionnent selon le même principe. Il n’existe pas d’appareil filtrant, et le mécanisme primaire est un transport actif d’ions (ions bivalents chez les Poissons, potassium chez les Insectes), qui entraîne un flux d’eau passif. La composition de l’urine primaire est alors très différente de celle du plasma ou de l’hémolymphe (elle est ainsi très enrichie en potassium chez les Insectes).Dans l’ensemble des cas considérés, l’urine éliminée est très différente de l’urine primaire. Cette modification est liée à des processus de sécrétion (NH3, par exemple) et de réabsorption tubulaires ou se produisant dans des parties extra-rénales: vessie des Amphibiens, cloaque des Oiseaux, rectum des Insectes...Protonéphridies et métanéphridiesLes protonéphridies apparaissent chez les Platheminthes (planaires). Elles dérivent dans ce cas de l’ectoderme, mais cela n’est pas une règle générale. Elles sont constituées par un ensemble de cellules qui s’ouvrent sur un canal excréteur. Cette structure peut être unique, paire ou multiple, à disposition métamérique (Annélides). Les protonéphridies, comme de nombreux organes excréteurs, ont un double rôle: élimination des déchets du métabolisme et régulation osmotique-ionique. La vitesse de filtration (de formation de l’urine primaire) est directement liée à la fréquence des battements de la flamme ou des flagelles.Les métanéphridies sont d’origine mésodermique. Elles sont ouvertes à leurs deux extrémités. L’ouverture interne, dans le cœlome (néphridiostome, ou pavillon cilié), réalise une filtration et non une ultrafiltration (celle-ci se produit entre le système circulatoire et le cœlome). Ce système est anatomiquement très comparable aux néphrons ouverts des embryons des Vertébrés, qui possèdent des glomérules intracœlomiques. La structure des métanéphridies est en fait complexe, et le tubule comporte plusieurs régions spécialisées dans la sécrétion ou la réabsorption (fig. 6). Ces échanges se produisent avec des vaisseaux sanguins qui longent le tubule.Chez les Annélides, les métanéphridies sont nombreuses: une paire par métamère. Chez les Mollusques, au contraire, le cœlome est réduit au cœlome péricardique et périgonadique, et l’on trouve soit une paire de métanéphridies ouvrant dans le péricarde, soit même une seule métanéphridie, comme dans le cas des Gastéropodes pulmonés où elle porte le nom d’«organe de Bojanus». La structure en est complexe (fig. 7). L’urine primaire se forme par ultrafiltration au niveau de la paroi cardiaque et de la partie proximale de la néphridie, richement vascularisée, qui porte le nom de «rein». L’épithélium de ce rein est très plissé, et sécrète en outre des grains de concrétions riches en urates et en diverses purines. L’urine finale est hypo-osmotique, en raison d’une importante réabsorption de sels dans la partie distale de la néphridie (uretère). On voit là de grandes analogies avec les Vertébrés, analogies qui sont soulignées par l’emploi des mêmes termes (rein, uretère). Chez les Mollusques céphalopodes (marins), l’épithélium néphridien sécrète des ions ammonium et non des urates, mais, malgré une anatomie quelque peu différente, le fonctionnement est très comparable.Chez les Crustacés (écrevisse), on trouve une paire de reins céphaliques. Il s’agit, là encore, de métanéphridies où la réduction du cœlome rapproche considérablement le vaisseau sanguin afférent (artère antennaire) de la partie proximale de la néphridie, ce qui conduit à une structure analogue à celle d’un néphron glomérulé.Les tubes de Malpighi des InsectesL’appareil excréteur des Insectes comporte un nombre variable de tubes excréteurs, ou tubes de Malpighi, qui sont aveugles à leur extrémité distale et qui débouchent à la limite entre l’intestin moyen et l’intestin postérieur. Les tubes de Malpighi baignent dans l’hémolymphe. La formation de l’urine est liée à un transport actif d’ions potassium par les cellules de la partie distale: il n’y a ici ni filtration, ni ultrafiltration. Le flux d’ions potassium entraîne un flux d’eau passif. Diverses substances, présentes en solution dans l’hémolymphe, diffusent ensuite passivement vers la lumière du tubule, selon le gradient de concentration. Certaines d’entre elles, en particulier l’acide urique, pourraient faire l’objet d’un transport actif. L’urine primitive ainsi formée est sensiblement iso-osmotique à l’hémolymphe, bien que sa composition soit notablement différente, car elle est très enrichie en ions potassium. Elle est neutre ou légèrement alcaline, l’acide urique y est présent sous forme d’urate de potassium, relativement soluble.L’appareil excréteur est en fait constitué par l’ensemble tubes de Malpighi + intestin postérieur (iléon + rectum). Au niveau de l’intestin, l’urine est acidifiée, ce qui précipite les urates sous la forme d’acide urique; les ions et l’eau sont en grande partie réabsorbés, de même que de petites molécules organiques solubles. En fait, chez de nombreuses espèces, la réabsorption commence dans la partie proximale des tubes de Malpighi. Le système excréteur peut présenter diverses adaptations, permettant en particulier une plus grande efficacité dans la réabsorption de l’eau (système cryptonéphridien des chenilles de Lépidoptères, par exemple).Les néphrons des VertébrésLes reins des Vertébrés sont constitués de néphrons (fig. 4 a), qui comportent en général deux parties: le corpuscule de Malpighi (ensemble glomérule + capsule de Bowman) et le tubule, où s’effectuent réabsorptions et sécrétions. Ce schéma correspond à la majorité des Vertébrés adultes, mais on connaît deux variantes, la première correspondant aux néphrons ouverts , analogues à des métanéphridies, que l’on rencontre au cours du développement embryonnaire des Vertébrés inférieurs, la seconde aux néphrons aglomérulés de certaines espèces de Poissons téléostéens marins, qui fonctionnent par sécrétion (transport actif d’ions bivalents Mg++, Ca++, S4—, HP4—), à la manière des tubes de Malpighi des Insectes (fig. 4 c). Le tubule comprend une partie proximale (réabsorption obligatoire, sécrétion) et une partie distale (réabsorption facultative modulable), qui sont séparées par un segment intermédiaire, essentiellement développé chez les Oiseaux et surtout les Mammifères, où il est recourbé en U et forme l’anse de Henlé , à laquelle est liée l’aptitude à produire une urine hyperosmotique. Chez les autres Vertébrés, l’urine est soit iso-osmotique, soit hypo-osmotique.Les reins sont formés d’un nombre très variable de néphrons selon les espèces: de quelques dizaines à quelques millions – deux millions chez l’Homme. Leur mise en place s’effectue au cours du développement embryonnaire, en concernant des métamères de plus en plus distaux. On note successivement la formation d’un pronéphros , en position antérieure, qui n’est fonctionnel que chez les embryons des Vertébrés inférieurs, puis d’un mésonéphros , qui forme le rein des Poissons et des Amphibiens, enfin d’un métanéphros , qui constitue le rein des Vertébrés amniotes (Reptiles, Oiseaux et Mammifères).L’urine émise par les reins transite par les uretères et est souvent stockée transitoirement dans une vessie. Chez les Poissons téléostéens, les Amphibiens et les Reptiles chéloniens, cet organe peut jouer un rôle important, en réabsorbant de l’eau et diverses substances dissoutes. Chez les autres Reptiles et chez les Oiseaux, les voies urinaires rejoignent les voies digestives au niveau du cloaque, et l’urine remonte dans le colon. Dans ce cas, la muqueuse intestinale peut réabsorber l’eau et les sels, modifier le pH et accroître la précipitation de l’acide urique à la périphérie des fèces, ce qui leur donne leur couleur blanchâtre. Ce dernier cas est fonctionnellement très proche de l’ensemble tubes de Malpighi + rectum des Insectes.Les Mammifères, grâce au développement de l’anse de Henlé qui permet la formation d’une urine hyper-osmotique, possèdent la faculté de concentrer l’urine plus ou moins fortement selon les espèces, cela étant en relation avec le mode de vie: les espèces marines (baleines, phoques), ainsi que celles vivant en milieu désertique (rat-kangourou, dromadaire) présentent une aptitude maximale à cette fonction, liée à la très grande longueur des anses de Henlé (cf. vie dans les DÉSERTS).Le problème de l’élimination des sels en excès se pose également chez d’autres Vertébrés vivant en milieu marin ou désertique (Oiseaux, comme les goélands, Reptiles: iguanes, tortues, etc.), qui ne possèdent pas cette aptitude à concentrer l’urine (absence ou faible développement de l’anse de Henlé). Dans ce cas, les sels en excès sont éliminés par des organes spécialisés, les glandes à sels, capables de sécréter des solutions concentrées de chlorure de sodium ou de potassium.Les activités rénales sont contrôlées à divers niveaux, celui de la filtration glomérulaire, celui des transferts tubulaires. Il faut toutefois noter que ces contrôles sont plutôt à mettre en relation avec les problèmes d’osmorégulation que ceux d’excrétion (cf. OSMORÉGULATION, métabolisme HYDROMINÉRAL).Quelques aspects évolutifs de la fonction excrétriceAu cours de l’évolution animale, on rencontre successivement des protonéphridies, puis des métanéphridies et, enfin, des néphrons glomérulés, dont nous avons vu les similitudes de fonctionnement.Cette évolution se retrouve au cours du développement embryonnaire ou larvaire: ainsi, les larves trocophores des Mollusques possèdent des protonéphridies à solénocytes ou à cirtocytes, les adultes des métanéphridies; les embryons de Poissons possèdent des néphrons ouverts (équivalant à des métanéphridies), les adultes des néphrons fermés classiques.Des systèmes homologues peuvent présenter des adaptations convergentes: les larves de certains Mollusques marins possèdent des protonéphridies dépourvues de solénocytes ou cirtocytes, ce qui peut être rapproché du cas des néphrons aglomérulés de certains téléostéens marins.Les similitudes physiologiques s’accompagnent de similitudes biochimiques concernant la nature des principaux déchets azotés, qui ne dépend pas de la position systématique des animaux, mais du milieu dans lequel ils vivent. Les produits terminaux du métabolisme azoté ne sont pas nécessairement uniquement des déchets. Outre un rôle dans la pigmentation, signalé plus haut, ils peuvent avoir un rôle dans l’osmorégulation. C’est ainsi que l’urée, en s’accumulant dans le sang (et les cellules) sert à rendre les Sélaciens iso-osmotiques à l’eau de mer, ou à empêcher la dessication des Dipneustes et des Amphibiens lors de périodes d’estivation pendant lesquelles ils sont enfouis dans de la boue (dont la pression osmotique s’accroît au cours de l’assèchement). Ce même composé contribue également à l’économie de l’eau chez les Mammifères, en participant aux mécanismes de concentration urinaire.
Encyclopédie Universelle. 2012.